
作者:解螺旋.蛋炒饭 解螺旋原创
新陈代谢简单的概括起来是合成代谢和分解代谢。以一个细胞水平去理解的话,代谢就是从细胞外摄取并利用营养,然后排出利用后的残渣,以此维持细胞生命活动的有序进行,这实际上是一种高度组织的熵减过程,是生命的基本特征。
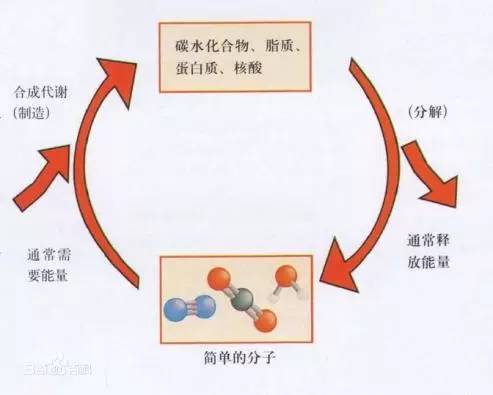
生命活动需要能量,细胞也需要一些生长以及代谢所必须的相关结构和蛋白,于是也需要一些简单的小分子来合成所需物质,能量和这些小分子都是碳水化合物的分解所得。能量的代谢通常都伴随着物质的代谢,物质循环也伴随着能量的产生与利用,这样的过程需要根据细胞与机体的物质和能量的需求状态进行精细的调节以维持生命活动的稳定。而明星分子mTOR则是调节生长与代谢的这样的一个枢纽分子。
为什么细胞信号转导通路里面mTOR的分子量如此之大?这主要由mTOR调控的过程所决定:
首先mTOR既然作为物质能量代谢调节的枢纽,本身不能由转录所调控,细胞应激中通过转录产生的蛋白分子量不会很大。生物进化中,如果大分子量的蛋白参与应激反应,其合成分解相对来说是一种较大的浪费。
再者,既然物质能量在细胞的各项过程中总在微妙的调节平衡中,那么这样的一个调节中心逻辑上可以应对多种的细胞环境或细胞需求或细胞应激,从而受到各种不同来源的调节,而大分子量蛋白相对来说显然更适合一些。而且从mTORC1和mTORC2之间的动态平衡也可以理解这一点(可参阅上期内容,回复:好基友查看)。
给大家一个临床上面的问题开放思考:rapamycin理论上不结合mTORC2而直接结合mTORC1抑制其活性,一般情况下只会抑制mTORC1的活性,而临床上发现长期的rapamycin则也会抑制mTORC2的活性,这是为什么呢?虽然目前科学家们并没有对这个问题给出明确的解释,但我们科研工作者也可以根据对信号的生命的理解,培养自己的生命的逻辑思维给出自己的解释。
下面几个方面跟大家讨论下mTOR与代谢的调节的分子机制:
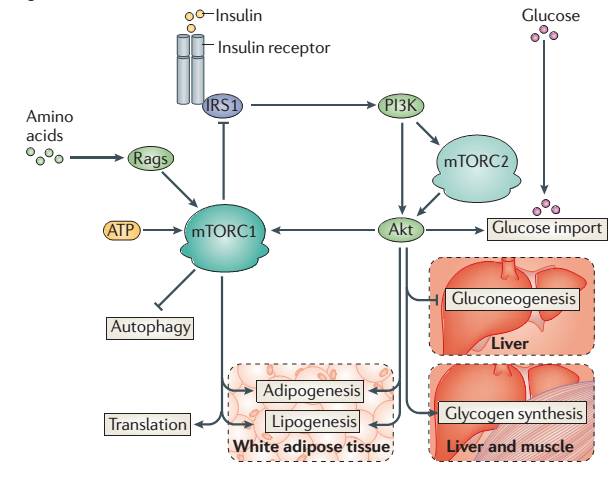
1.对胰岛素和胰岛素样生长因子的响应
PI3K-AKT-mTOR是经典的响应胰岛素信号的通路。当进食后,被分解的葡萄糖进入血液促进胰岛素的释放,胰岛素作为响应营养富余的信号,会指导细胞进行吸收利用这些营养。Insulin首先结合细胞表面受体通过IRS1激活PI3K-AKT通路,AKT直接促进Glucose的吸收,同时通过AKT-TSC1/2-RheB-mTORC1激活mTORC1的活性,mTORC1进一步指导合成利用glucose进行生物合成相关的酶进行营养的储存。RheB是一种小G蛋白,小G蛋白激活后可直接调控mTORC1,介导的激素类即细胞表面信号到mTORC1,这是众多mTORC1调节方式中的一种方式。
这一过程既包括吸收也包括吸收后的利用调节,这样一条龙的调节方式,通常被很多科研工作者理解为mTORC1被激活后,既可以合成糖原、可以促进翻译,也可以促进生长。这样笼统的理解显然是有失偏颇的,也是我们经常对文献理解的一个误区。实际上Insulin激活mTORC1大部分情况下是和糖代谢相关,而IGF(胰岛素样生长因子)激活mTORC1才主要涉及细胞的生长相关。mTORC1激活后分别调控的底物理论上是有差别的,非常具体的调控与底物信息等仍有待进一步研究。
2.对氨基酸-营养物质的响应
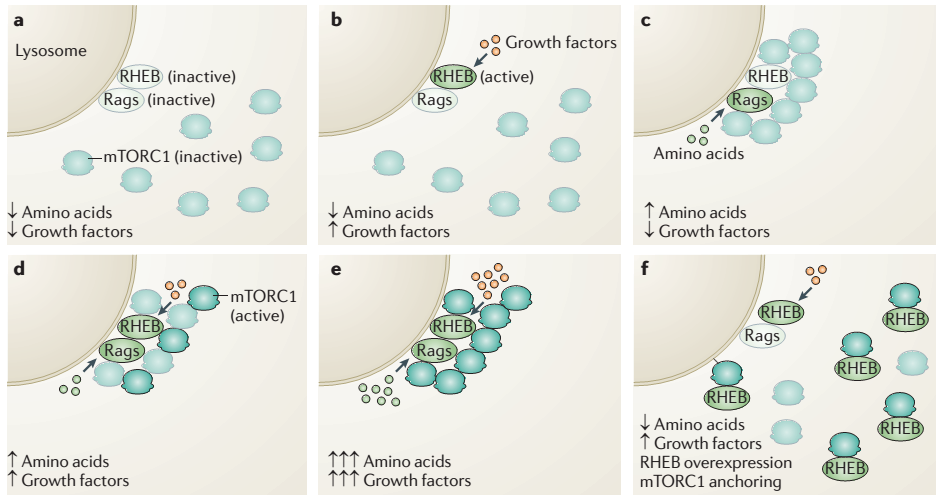
三大营养物质其中糖类,前面已经提到主要通过Insulin与细胞表面受体间接调节mTORC1的活性。而氨基酸被吸收后则可以直接激活mTORC1的活性。Sabatini发现mTORC1的激活主要由2步:首先在氨基酸存在的情况下,小G蛋白Rags被激活,结合mTOR协助其定位于溶酶体膜表面。然后定位于膜表面的小G蛋白Rheb蛋白被来自生长因子的信号激活后才可以激活mTOR的活性。这里的由氨基酸直接激活mTOR的调节模式虽然不经由来源于细胞表面的信号,也是由小G蛋白Rags所介导。
既然在growth factor和氨基酸都存在的情况下mTOR才可以被完全激活,那也就是在物质和能量都富余的情况下才具备活化mTOR的条件。这一点正说明了mTOR作为代谢调节的枢纽具有信息整合的作用。试想一下,如果glucose富余而氨基酸短缺的情况下,细胞拿什么材料去合成,来完成生长呢?所以这个时候glucose更多的是用来转化成氨基酸而非能量储存,所以mTOR未被激活。相反在氨基酸富余而glucose不足的情况下,合成所需的能量补给不足,氨基酸这个时候更多的可能会被用来分解而提供细胞活动所需能量,所以mTOR也必然不能被激活。这正看出mTOR的调节模式也反映了细胞代谢平衡的机智的策略。比较有意思的是,两条线路都是经由小G蛋白的信号,那么细胞的代谢稳态比如氨基酸和糖类之间的生化转化的信号如何感知呢?是否小G蛋白参与其中起到重要的作用呢?引人遐想。
2015-08-12