
作者:解螺旋.墨斗鱼
俗话说“一个好汉三个帮”,TGF-β作为信号通路中的明星人物没几个帮手又怎么能完成信号转导这个艰巨的任务呢!要说在这些明星中谁才是与TGF-β的最佳好基友,那非Smad家族莫属。所以要了解TGF-β信号通路,我们今天先把Smad家族一探究竟。

一.Smad信号转导通路
Smad蛋白(Smad是由线虫的Sma和果蝇的Mad缩写而来)是TGF-β家族受体下游的信号转导分子[1,2]。在哺乳动物中有八个Smad蛋白, Smad1、 Smad1、 ...、 Smad8。
 >
>
根据在TGF-β信号转导中的功能差异,它们被分为三类:
-
受体调控的Smad(receptor-regulatedSmad,R-Smad–Smad1/2/3/5/8)
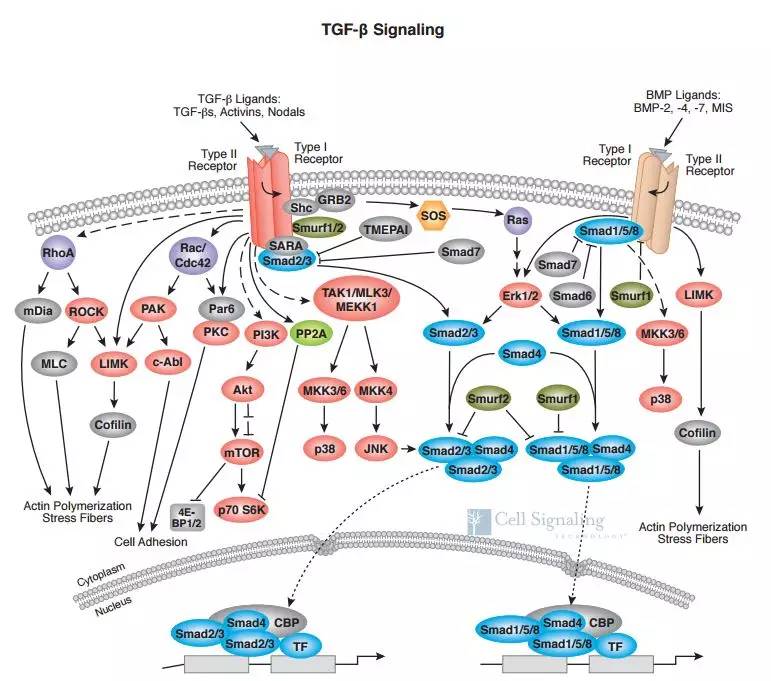
与其它跟许多激酶-底物反应一样,TGF-βI型受体对R-Smad的磷酸化需要它们的直接结合。R-SmadC末端SXS基序(X指代M或V)中的丝氨酸(S)能直接被TGF-βI型受体磷酸化而导致R-Smad活化,其中Smad2/3被TGF-β/activin/nodal亚家族的I型受体ALK4/5/7磷酸化,而Smad1/5/8则主要被BMP/GDF/AMH亚家族的ALK2/3/6和ALK1磷酸化[2]。
-
通用Smad(common Smad,Co-Smad–Smad4)
磷酸化的R-Smad与Co-Smad/Smad4聚合并继续传递信号。R-Smad与Smad4的聚合主要以异源三聚体的形式存在,并以二聚体的形式发挥作用,这可能与基因启动子区的结构有关。最终导致R-Smad-Smad4复合物在细胞核内的积累并调控靶基因转录[3,4]。
-
抑制型Smad(inhibitorySmad,I-Smad –Smad6/7)
抑制型Smad主要起抑制信号转导的作用。I-Smad(包括Smad6和Smad7)是TGF-β信号的关键负调控因子;它们本身是TGF-β/BMP的靶基因,因此形成负反馈的调控模式[5]。Smad7是TGF-β家族的广谱抑制蛋白,而Smad6更特异性的抑制BMP信号,尤其是ALK3/6介导的信号。
二.非Smad信号转导通路
很早以前科学家们就发现TGF-β/BMP细胞因子能在多种细胞中激活一些非Smad信号分子。比如TGF-β活化的TGF-βRI通过磷酸化ShcA而诱导形成ShcA/Grb2/Sos复合物,并进一步通过Ras-Raf-MEK通路激活ERK MAPK[6];TGF-β受体诱导TRAF6催化TAK1发生K63连接的多聚泛素化修饰,然后通过MKK激活JNK和p38 MAPK[7];BMP也能通过TAK1/TAB激活MAPK通路[8]。
TGF-β还能诱导其受体与PI3K的调节亚基p85结合,激活PI3K及其下游的Akt或者PAK2激酶[9];非受体酪氨酸激酶FAK能与p85结合并控制Akt/PAK2的活化[10]。此外,TGF-β能调控RhoA、LIMK1/2、PP2A、eEF1A1等信号分子活性[4]。TGF-β诱导的非Smad通路通常依赖于不同的生理/环境条件和细胞类型,并与Smad通路一起介导TGF-β诱导的生物学反应,比如上皮细胞-间充质细胞转化(epithelial-mesenchymal transition, EMT)、肿瘤细胞的迁移与侵袭、成纤维细胞的增殖与形态改变,以及组织纤维化等。
TGF-β诱导的非Smad通路能够对Smad信号进行调控。ERK、JNK和p38 MAPK能够磷酸化R-Smad和Smad4,尤其是它们的linker区。这种磷酸化既可能促进,也可能抑制Smad的活性,这依赖于细胞类型以及细胞所处的生理环境[11]。Akt直接结合Smad3并抑制它被受体的磷酸化、入核转运和转录活性[12]。最近我们发现在MDCK等表皮细胞中,PAK2通过磷酸化Smad2(以及Smad3)而抑制其活性[13]。
最后是终结性的TGF-β信号通路图,基本涵盖了TGF-β信号通路的大部分内容。希望给大家帮助。在下一次的TGF-β信号通路介绍中,我将会更细致的内容带领大家梳理TGF-β的精细调控。
参考文献:
[1] Feng,X. H. , et al. (2005). Specificity and versatility in tgf-beta signalingthrough Smads. Annu Rev Cell Dev Biol 21, 659-93.
[2] Heldin,C. H., et al. (1997). TGF-beta signalling from cell membrane to nucleus throughSMAD proteins. Nature 390, 465-71.
[3] Massague,J., et al. (2005). Smad transcription factors. Genes Dev 19, 2783-810.
[4] Moustakas,A. et al. (2009). The regulation of TGFbeta signal transduction. Development136, 3699-714.
[5] Lonn,P., et al. (2009). Regulating the stability of TGFbeta receptors and Smads.Cell Res 19, 21-35.
[6] Lee,M. K., et al. (2007). TGF-beta activates Erk MAP kinase signalling throughdirect phosphorylation of ShcA. EMBO J 26, 3957-67.
[7] Yamashita,M., et al. (2008). TRAF6 mediates Smad-independent activation of JNK and p38 byTGF-beta. Mol Cell 31, 918-24.
[8] Bragdon,B., et al. (2011). Bone morphogenetic proteins: a critical review. Cell Signal23, 609-20.
[9] Wilkes,et al. (2003). Cell-type-specific activation of PAK2 by transforming growthfactor beta independent of Smad2 and Smad3. Mol Cell Biol 23, 8878-89.
[10] Hong,M., et al. (2011). Non-Smad transforming growth factor-beta signaling regulatedby focal adhesion kinase binding the p85 subunit of phosphatidylinositol3-kinase. J Biol Chem 286, 17841-50.
[11] Wrighton,K. H., et al. (2009). Phospho-control of TGF-beta superfamily signaling. CellRes 19, 8-20.
[12] Conery,A. R., et al. (2004). Akt interacts directly with Smad3 to regulate thesensitivity to TGF-beta induced apoptosis. Nat Cell Biol 6, 366-72.
[13] Yan,X., et al. (2012). p21-Activated kinase 2 (PAK2) inhibits TGF-beta signaling inMadin-Darby canine kidney (MDCK) epithelial cells by interfering with thereceptor-Smad interaction. J Biol Chem 287, 13705-12.
2015-08-03