
作者:解螺旋.大河马
从 Hippo 信号通路被确定以来,因为其在器官大小和肿瘤发生调节中的重要作用(回复394,查看相关内容),研究者们一直致力于发掘新的 Hippo 信号通路的调节因子。其中,核心激酶链蛋白调控 Hippo信号通路是人们研究的重点之一。
2006年,Hamaratoglu等人发现两个FERM (4.1, Ezrin, Radixin, Moesin) domain-containing family 蛋白—Merlin (Mer)和 Expanded(Ex)为 Hpo 上游的调节因子。
Mer 和 Ex 能相互结合在上皮细胞的顶层,它们与后来发现的 Kibra 共同作用刺激 Hippo 信号通路。Kibra,Ex 和 Mer 能协同促进 Wts 的磷酸化;Kibra,Ex 和 Mer 复合体能直接和 Hpo-Sav 复合体结合,对于 Hpo 在细胞膜的定位非常重要。这提供了一种可能性,那就是 Hpo 信号通路的核心组分被募集到细胞膜上能激活 Hpo信号通路的活性。另外,DJ Pan 教授课题组发现 Mer 能够直接将 Wts 募集到细胞膜上,经 Hpo-Sav 复合体磷酸化后激活。这项研究进一步证实了这种可能性。
细胞连接与顶层-基底层极性复合物对 Hpo 信号通路的调控
跨膜蛋白 Crb(Crumbs)是调控顶层-基底层极性的重要蛋白。Crb 通过FBM 区域与 Ex 结合并影响 Ex 的亚细胞定位及稳定性,进而影响下游信号。另外,调节上皮细胞顶层-基底层极性的复合物 aPKC(Bazooka、Par6 和 aPKC)以及 Scrib 也参与了 Hippo 信号通路的调控。比如 aPKC 能促进 Yki 的活化,Scrib通过拮抗 aPKC 复合物活性抑制 Yki 的活性等。
上皮细胞之间由细胞连接黏附在一起,最为典型的细胞连接包括紧密连接(tight junctions,TJs)和粘着连接(adheres junctions,AJs)。
Angiomotin(AMOT)家族蛋白包括 AMOT、AMOTL1 和 AMOTL2,在维持 TJs 稳定性方面非常重要。研究发现,AMOT 能与 YAP 和 TAZ 直接相互作用,从而将其转移到细胞膜上,降低它们的活性。这种相互作用不依赖 YAP 和 TAZ 的磷酸化。另外,AMOT 家族成员可以通过与 Lats2 的相互作用激活 Lats2 从而抑制 YAP 的活性。Merlin/NF2 与 Angiomotin, Patj 和 Pals1 形成复合体从而被定位到 TJs 上。它与小 GTPase 激活蛋白 Rich1 竞争性结合 Angiomotin,从而抑制 Ras-MAPK信号通路。
还有众多对建立或维持细胞极性起重要作用同时又调控 Hpo信号通路的蛋白,例如:a-catenin、protein tyrosine phosphatase14(PTPN14,即果蝇中的 Pez)、E -cadherin、Ajuba、liver kinase B1(LKB1)、nephronop hthisis 4(NPHP4)、ZO-1/2 等等。
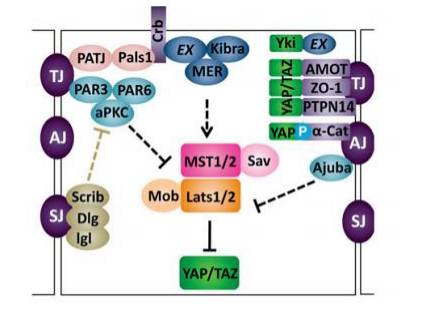
顶层-基底层极性对 Hpo 信号通路的调控(Yu F X, Guan K L . Genes Dev, 2013)
平面细胞极性对 Hpo 信号通路的调控
平面细胞极性(Planar Cell Polarity,PCP)也是调控 Hippo 信号通路的重要因素。Ft(Fat)是一种非常规的原钙黏蛋白,它含有多个钙黏素重复序列,是第一个被发现调控 Hippo 信号通路的膜蛋白,它既能调控细胞生长又能调控 PCP。
目前发现,Ft 调控 Hippo 信号通路可能通过两种机制:一种是 Ft, Ex 和 Hpo 是一种线性的关系,其中 Ft 能影响 Ex 的亚细胞定位和稳定性,进而调控 Hippo 信号通路;另一种模型就是 Ft 和 Ex 是平行的关系,它们分别都能调控 Wts 的磷酸化。其中 Ft 也能通过 D(Dachs)和Zyx(Zyxin)影响 Wts 的稳定性。Ds (Dachsous)是一种原钙黏蛋白,它能够与 Ft 结合促进其活性。另外,Ft 还受好多其他蛋白的调控,比如酪蛋白激酶 Dco (Discs overgrown), 高尔基体蛋白激酶 Fj (Four-joined),和 Lft (The Fat/Ds-interacting protein Lowfat)。
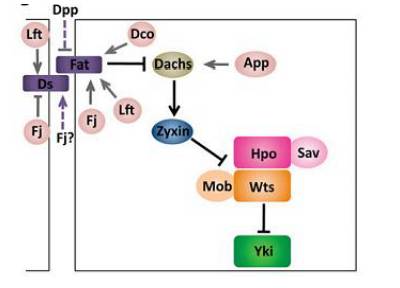
平面极性调控 Hippo 信号通路网络(Yu F X, Guan K L . Genes Dev, 2013)
细胞膜受体调控hippo信号通路
细胞膜上存在很多受体,它们能感受外界生长因子的刺激,从而激活细胞内相关信号通路调节细胞的增殖或凋亡。所以,Hippo 信号通路很可能也对某种生长因子刺激有反应。然而很多常见的生长因子比如胰岛素(insulin)和上皮生长因子(EGF)都对 YAP 的磷酸化没有显著影响。
两个独立的课题组发现,血清中的 LPA 和 S1P 能迅速激活体外培养的细胞中 YAP/TAZ 。两项工作都指出 LPA 和 S1P 能够结合它们在膜上的受体 GPCRs,通过 Rho GTP酶激活 YAP/TAZ。另外一项研究指出,凝血酶(thrombin)能激活蛋白酶激活受体(PARs,一种 GPCR)进而激活 YAP/TAZ。进一步的研究证明 YAP/TAZ可以受很多 GPCR 及其配体的激活,这说明 GPCR 作为一类膜受体在调节YAP/TAZ 的普遍性。Gα12/13, Gaq/11 或者 Gαi/o-coupled signals 能激活 YAP/TAZ,而 Gαs-coupled signals 能抑制 YAP/TAZ 的活性。
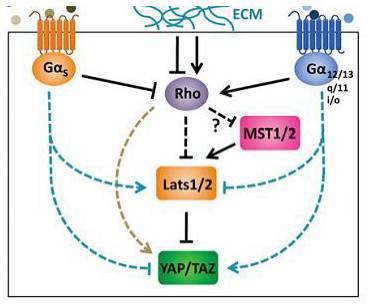
GPCR 信号和 ECM 调控 Hippo 信号通路(Yu F X, Guan K L . Genes Dev, 2013)
细胞骨架对 Hpo 信号通路的调控
体内细胞能感受到邻近细胞、细胞外基质、细胞周边体液传导过来的以及细胞自身形态改变所产生的机械力,并通过调控细胞的增殖、分化、凋亡或者迁移以适应机械力变化造成的影响。微丝与微管作为细胞骨架重要组成部分在感知和转导机械力信号方面起着重要作用,并参与了对细胞增殖与分化的调控。最新研究发现,在果蝇及哺乳动物细胞中微丝均可以通过 Hpo 信号通路调控细胞的增殖。
2015-07-30