作者:解螺旋.大河马
大象如此之大,老鼠如此之小?

双手双脚为什么能发育的如此相似?

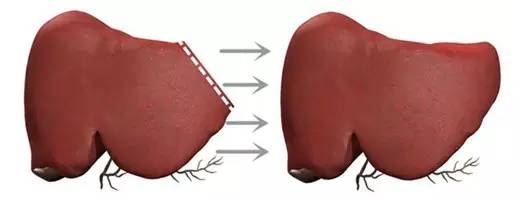
为什么肝脏在切除2/3之后还能发育?
······
这些千奇百怪的问题,无非是因为人类在发展长流中,总是在试图弄清楚:控制个体大小的机制是什么?(人类解决了温饱之后,开始对科学世界、精神领域产生了浓厚的追求)Hippo信号通路的发现也许为其提供很好的解答切入点。

组织器官的发育不仅要塑造成型(patterning),还要有其形(growth)。只有这两个过程都经历了,眼睛才是眼睛,鼻子才是鼻子!关于发育遗传的机制研究中,过去几十年(2002年之前)至少揭示了7条信号通路,包括:Notch、Wnt、TGF-、Hedgehog、receptor tyrosine kinase、nuclear receptor and Jak/STAT。
而在之后,科学家开始关注发育中组织器官发育的大小调控机制,比如the tuberous sclerosis tumor suppressor complex (TSC)–target of the rapamycin (TOR) pathway:通过调控细胞生长(growth)来控制组织大小。Hippo信号通路通过调控细胞生长、增殖、凋亡来控制组织大小。

先来看看Hippo信号通路的pathway吧,如下图,a为在果蝇中的通路,b为在哺乳动物中的通路图。
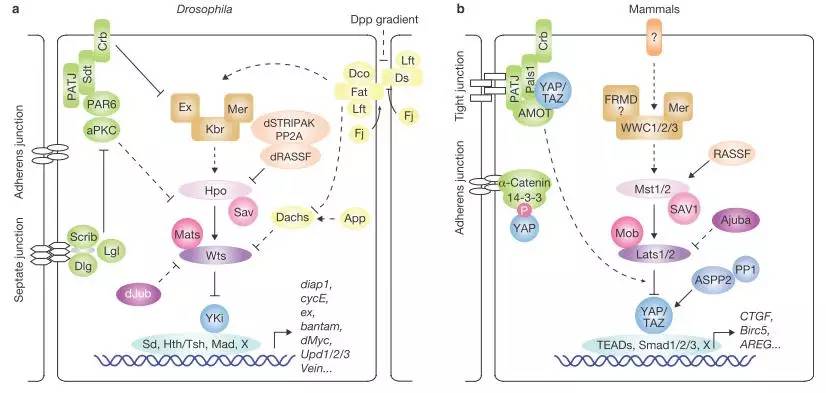
Bin Zhao et al.,2013, NATURE CELL BIOLOGY
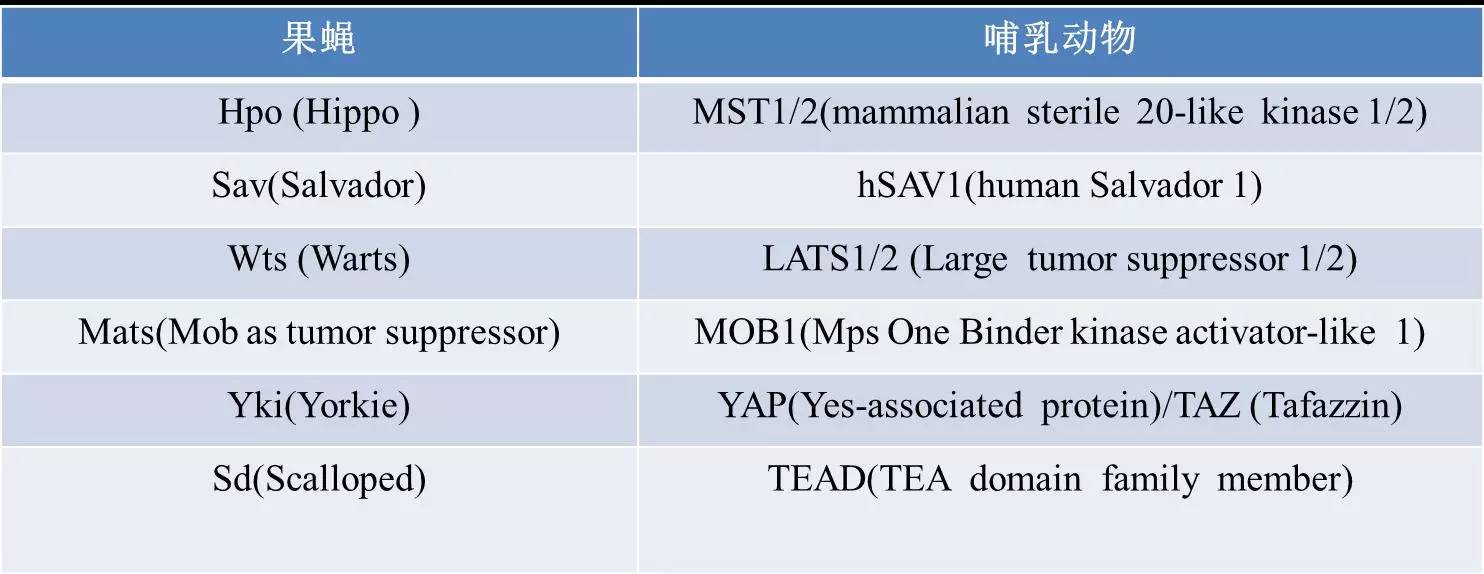
Hippo信号通路在进化过程中高度保守的,果蝇和哺乳动物中核心元件同源。如下表:
好了,梗概了解了,就步入正题说说他们的发现历史吧(下面专业姿势来了,请好好学习!)
先前的研究中显示,传统的遗传学筛选技术在果蝇中的利用具有较大的局限性,主要是基于纯合突变体,所有的必须基因在得到纯合突变体的时候导致果蝇的早期死亡,后续的功能验证分析受到阻碍。FRT/FLP 技术以及基于 FRT/FLP 技术发展而来的嵌合体技术在果蝇中的应用使得筛选肿瘤抑制基因变得更加便利高效。
1995 年,FRT/FLP 系统创始人 Xu Tian研究小组及 Justice,R.W. 研究小组同时利用嵌合体筛选技术发现了一个新的肿瘤抑制基因,分别命名为Lats (large tumor suppressor gene)和Warts。Lats/Warts 编码核 Dbf-2 相关家族(Nuclear Dbf-2 related family ,NDR)的丝氨酸/苏氨酸激酶。
在果蝇翅膀、腿和眼睛等诸多器官的上皮组织中,Wts 的功能丧失将导致细胞自主性的过度增生,但是不影响细胞的分化。Wts 作为肿瘤抑制基因,其发挥作用的分子机制模糊不清,其结合蛋白及上游和下游的调控因子也不得而知,这种“孤儿基因”的局面持续了 7 年,直到另一个肿瘤抑制基因 Sav 的发现。
2002年,Kango-Singh,M.和 Tapon,N.同时发现了一个新的基因,其突变后也会引起突变组织中细胞自主性的过度增生,类似于 Wts 突变的表型。 Kango-Singh,M.将其命名为 Shar-pei(Shrp),因为突变个体头部表皮过度增殖而引起的扭曲皱褶的形态类似于沙皮狗;而 Tapon,N.将其命名为 Sav。Sav 编码一个具有WW 结构域的蛋白,遗传学上位性实验表明 Sav 位于 Wts 的上游。Tapon,N.科研小组发现在 Wts 和 Sav 突变后促进细胞增值的同时抑制了细胞凋亡,首次证实两者协同调控细胞增值与凋亡。Tapon,N.等进一步发现,Wts 和 Sav 突变后会引起细胞周期调控因子 CyclinE(CycE)和细胞死亡抑制因子 Diap1 表达水平的上调,这表明 CycE 和 Diap1 可能是 Wts 和 Sav 的靶基因。肿瘤抑制基因 Hpo的发现是 Hpo 信号通路发展过程中的重大进展。Hpo 编码一个 Ste-20 家族蛋白激酶,其突变后引起的组织过度增生类似于 Sav 或 Wts 突变后的表型。Wu Shian 等首次阐释了 Hpo 激酶级联反应链,在这个激酶级联反应链中,Hpo 磷酸化并激活 Sav 和 Wts,Sav 与 Hpo 结合后促进 Hpo 对 Wts 的磷酸化。Wu Shian等进一步证实 Hpo 激酶级联反应链在转录水平对 Diap1 进行调控,这在后来的研究结果中一再被证实。Lai Zhichun 等在 2005 年发现果蝇中 Mob-1 相关蛋白Mats(Mob as tumor supressor)不仅能够结合 Wts,而且能促进 Wts 激酶活性;另外,Mats 与 Hpo 结合后被磷酸化,磷酸化的 Mats 进一步促进 Mats 和 Wts的结合。Mats 突变后与 Hpo、Sav 或 Wts 的突变表型类似。

Wu Shian et al., 2003.,cell
既然 Hpo 信号通路调控 Diap1 等的转录,那么一个重要的问题是必然存在这样一个转录因子,它受 Wts 激酶活性的调控同时又调节 Diap1 的转录。Huang Jianbin 等通过酵母双杂交技术发现转录辅因子Yki 是Wts 的底物和效应因子。Wts 磷酸化 Yki 继而抑制 Yki 的活性,最终导致 Diap1 转录水平下调;在果蝇中过表达 Yki 将导致组织过度增生,相反的,其功能丧失将造成组织萎缩;遗传学上位性实验同样表明 Yki 位于 Wts 的下游。Yki 的发现进一步证实 Hpo 信号通路通过转录水平而不是转录后水平调控其靶基因。至此,从Hpo 到 Yki 的激酶级联反应链的建立标志着核心 Hpo 信号通路的形成。
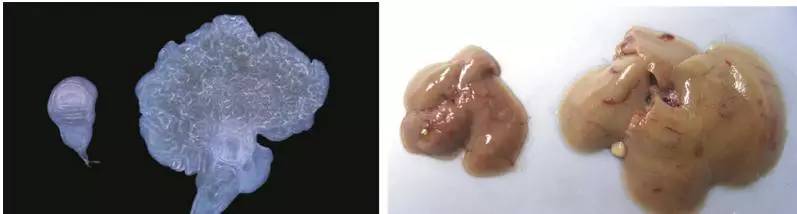
Huang et al., 2005,cell;
右为wt,左为过表达yki的翅膀成虫盘;右为wt,左为哺乳动物过表达YAP的肝脏。
这样就概括下,Hippo信号通路包括四大部分:
多重上游信号输入因子,包括 Fat4、Dchs1/2、FRMD6、NF2和KIBRA;
核心激酶级联反应链MST1/2、hSAV1、LATS1/2和MOB1;
下游转录共激活因子,包括 YAP/TAZ 和 TEAD;
以及下游调节因子,包括RASSFs、ASPP1/2和Ajuba。
2015-07-22